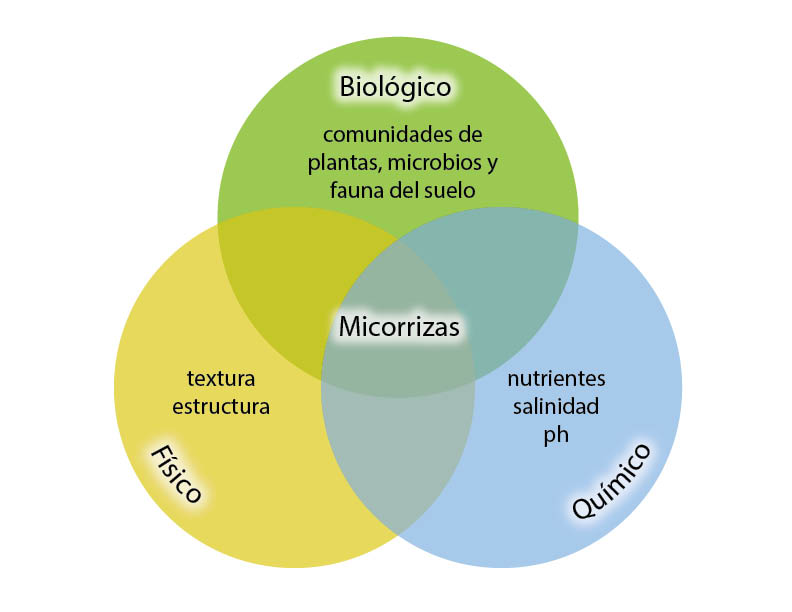
El suelo está compuesto de billones de microorganismos, incluyendo muchos miles de especies bacterianas y fúngicas, que uno puede encontrar en una sola cucharada de suelo (Delmont et al., 2014). En términos de investigación, la biodiversidad del suelo se ha convertido en una importante área de actividad, en diferentes niveles: taxonómico, genético o funcional. Entre los investigadores, la biodiversidad del suelo ha sido defendida por algunos como un factor crítico que controla la amplia gama de procesos microbianos que son cruciales para el funcionamiento adecuado de los suelos (Orgiazzi et al., 2015), regulan su capacidad para proporcionar servicios a poblaciones humanas (Nannipieri et al., 2003) y estabilizan las condiciones de vida en la tierra (Ferris & Tuomisto, 2015).
Se ha demostrado que la comunidad microbiana y su fisiología asociada es un fuerte impulsor en el desarrollo de fertilidad sugiriendo que la fisiología de la comunidad del suelo y sus entradas de residuos pueden ser tan importantes o más que los más efectos ampliamente reconocidos de la estructura mineral del suelo en la estabilización del Carbono (Kallenbach et al., 2016) donde los procesos microbianos de sustratos simples produjo una abundancia en la materia orgánica estable químicamente y diversa con alta presencia de proteínas y lípidos microbianos, que la hicieron comparable a suelos naturales, mejorando de este modo directamente la fertilidad.
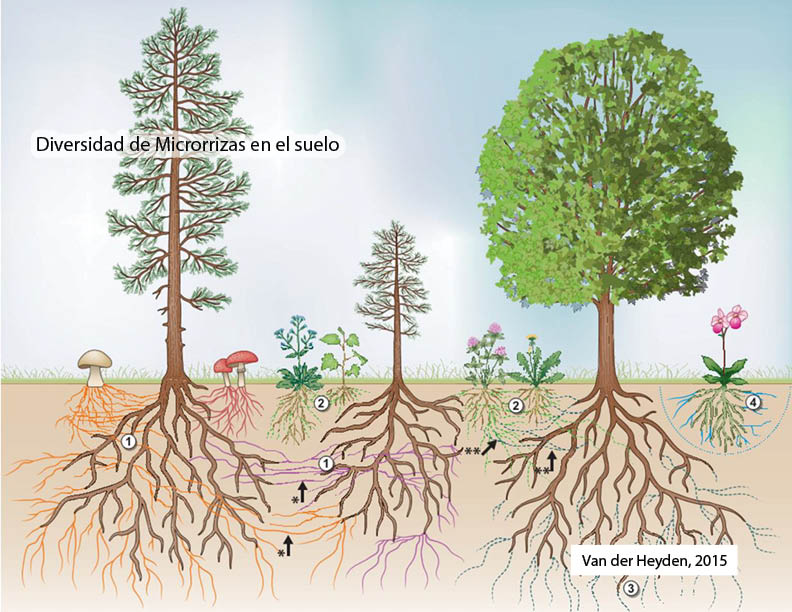
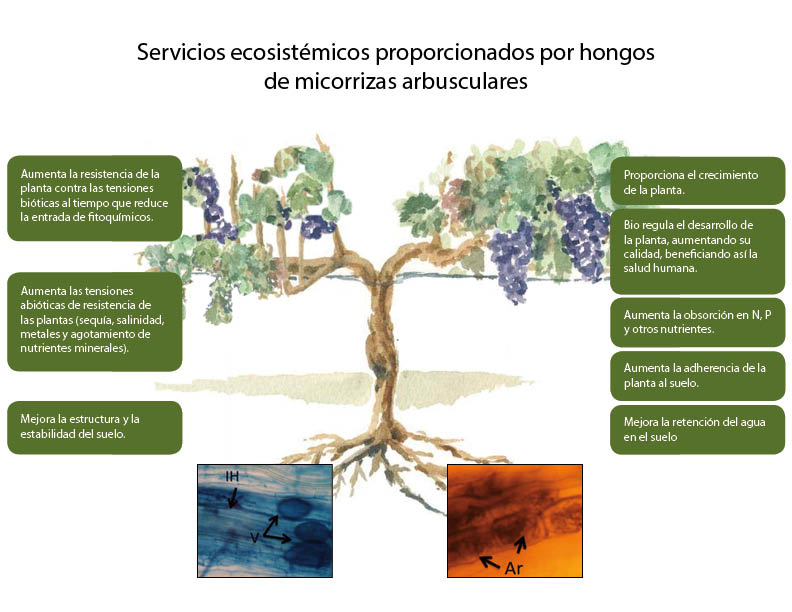
Los hongos micorrícicos arbusculares (HMA) forman una asociación simbiótica mutualista con las raíces de la mayoría de las plantas terrestres. En esta simbiosis la planta hospedera entrega azúcares provenientes de la fotosíntesis y el hongo deja a mayor disposición el agua y los nutrientes principalmente N y P. Los HMA son el tipo más común de micorriza, se encuentran asociadas al 80% de las plantas, existiendo hace más de 460 millones de años. La colonización con HMA otorga a la planta principalmente una mayor absorción de agua y macronutrientes como el N y el P, de baja disponibilidad en los suelos así como de algunos micronutrientes (Zn, Cu, etc.), lo que mejora su crecimiento y rendimiento. (Smith & Smith, 2012) Además permite una mayor resistencia a estrés hídrico, estrés salino y a presencia de metales pesados. La presencia de HMA, puede aumentar la tolerancia al déficit hídrico, pues las hifas pueden acceder a pequeños poros del suelo, aumentando el área de absorbancia de la raíz. (Worchel et al., 2013). Existe múltiple evidencia de la resistencia que generan ante el ataque de patógenos, desde bacterias, nematodos, hongos y algunos insectos que atacan la raíz (Jung et al., 2012). La formación de micorrizas arbusculares modifica la rizósfera, influencia la estructura del suelo y aumenta la capacidad exploratoria del suelo de la raíces en búsqueda de nutrientes y agua. Se cree que la mayor resistencia a patógenos está relacionada con la competencia por espacio y nutrientes en la raíz, así como por la inducción de cambios en el metabolismo primario y secundario y la regulación de los mecanismos de defensa de la planta, afectando también las partes aéreas de las plantas. (Toussaint, 2007). Esta capacidad de aumentar la tolerancia a distintos tipos de estrés biótico y abiótico, han permitido la sobrevivencia a través de años de evolución de esta simbiosis entre las plantas y los HMA (Worchel et al., 2013).
Ana Aguilar –Dra. Ciencias Biológicas
Bibliografía
Delmont, T.O., Francioli, D., Jacquesson, S., Laoudi, S., Mathieu, A., Nesme, J., Ceccherini, M.T., Nannipieri, P., Simonet, P., Vogel, T.M., 2014. Microbial community development and unseen diversity recovery in inoculated sterile soil. Biology and Fertility of Soils 50:1069-1076.
Ferris, H. & Tuomisto, H., 2015. Unearthing the role of biological diversity in soil health. Soil Biology & Biochemistry 85:101-109.
Jung S, Martínez-Medina A, Lopez-Raez J & Pozo M. 2012. Mycorrhiza-induced resistance and priming of plant defenses. Journal of Chemical Ecology 38(6):651-64.
Kallenbach, C. M. Serita D. Frey, A. Stuart Grandy. 2016. Direct evidence for microbial-derived soil organic matter formation and its ecophysiological controls. Nat. Commun. 7:13630 doi: 10.1038/ncomms13630.
Nannipieri, P., Ascher, J., Ceccherini, M.T., Landi, L., Pietramellara, G., Renella, G., 2003. Microbial diversity and soil functions. European Journal of Sport Science. Electronic Resource 54:655-670.
Orgiazzi, A., Dunbar, M.B., Panagos, P., de Groot, G.A., Lemanceau, P., 2015. Soil biodiversity and DNA barcodes: opportunities and challenges. Soil Biology & Biochemistry 80:244-250.
Smith S & Smith F. 2012. Fresh perspectives on the roles of arbuscular mycorrhizal fungi in plant nutrition and growth. Mycologia 104(1):1-13.
Toussaint J. 2007. Investigating physiological changes in the aerial parts of AM plants: what do we know and where should we be heading. Mycorrhiza 17(4):349-53.
Worchel E, Giauque H & Kivlin S. 2013. Fungal symbionts alter plant drought response. Microbiology Ecology 65(3):671-8.